Marchantia polymorpha - a new model system for Synthetic Biology
The liverworts (or Marchantiophyta) are descendants of the earliest terrestrial plants. The group is characterised by morphological simplicity, and this seems to be matched by simple underlying genome structures. Liverworts show promise as new experimental systems after recent developments in transformation methods and genome characterisation.
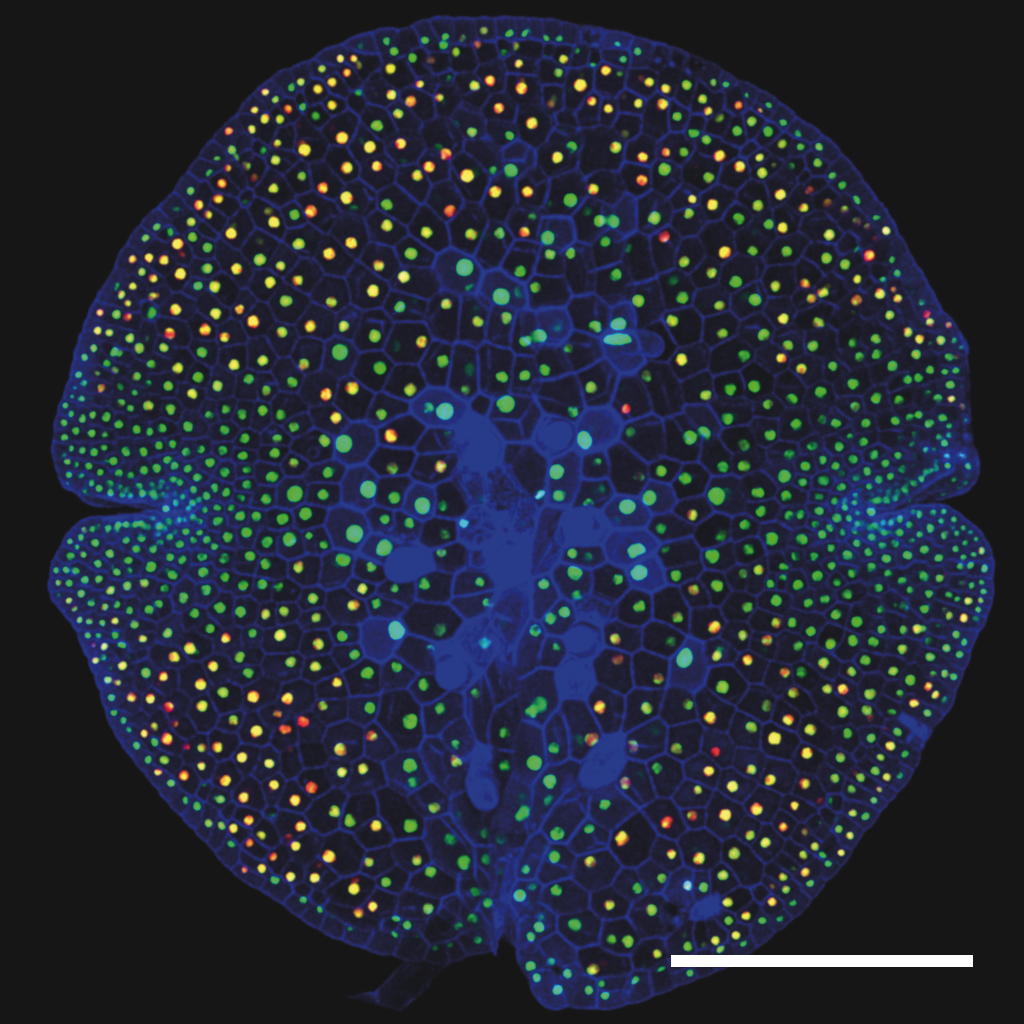
Marchantia polymorpha is the best characterised of the liverwort. It is a thalloid liverwort, growing as flat sheet-like tissues which possess distinct upper and lower surfaces. The lower surface of the plant has specialised root-like cells, or rhizoids. The body of the thallus contains oil bodies, present as scattered differentiated cells. This is characteristic of the liverwort group. The upper surface has a modular structure, with repeated formation of units that form primitive cell complexes adapted for photosynthesis and gas exchange. Each unit is marked by a single, permanently open central pore. The liverworts have less efficient systems for water transport and retention than higher plants, and can show marked tolerance to desiccation.
M. polymorpha plants are easy to grow. They show distinctly weed-like properties, growing vigorously on soil, artificial media such as vermiculite and rockwool, and sterile media.
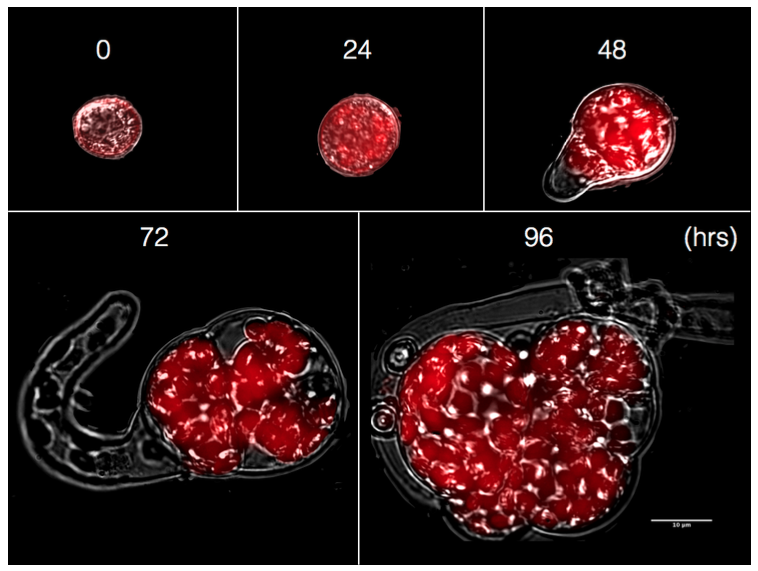
The liverworts have alternate haploid and diploid generations. Like other Bryophytes, the gametophyte or haploid generation is dominant phase of the life cycle. M. polymorpha has a global distribution, and is often found as a weed in horticulture. The plant produces vegetative propagules. These form vegetatively inside conical splash cups. Superficial cells on the inside of a cup undergo cell proliferation to form a group of cells carried on a short stalk, the cells continue to proliferate in regular fashion to form a bilobed gemma (see right). This is eventually detached from the stalk, and can be dispersed from the cup, typically by water splash. The propagules are robust and long-lived, even tolerating desiccation. Gemma can be used for simple vegetative propagation and amplification of plantlets during experiments.
Haploid liverwort plants are either male or female, with sex determined by presence of X or Y chromosomes. A PCR-based assay can be used to determine the sex of M. polymorpha plants. Gametogenesis is under environmental control, and can be induced by exposure to far-red light. Antheriophores and archegoniophores grow upward from the Marchantia thallus, with gamete-forming structures supported by pedestals. Antheridia form on the upper surface of a disc, and archegonia form underneath an arrangement of spokes. Genetic crosses can be performed by transfer of sperm from mature antheridiophores to archegonia by pipette. After fertilisation and zygote formation, the diploid phase of the life cycle continues with cell proliferation, meiosis and spore formation. The spores and elaters are packaged within yellow sporangiophores, suspended under archegoniphores. The elater cells are thickened spiral-shape cells which are hydroscopic and change their helical pitch. These aid the release of spores. Spores can be stored in a cool desiccated state for over a year.
Similar to other lower plants, M. polymorpha regenerates easily, and was used in many early tissue culture studies. A number of plastid and nuclear transformation techniques have been developed for M. polymorpha, including a recent method for Agrobacterium tumefaciens mediated transformation using germinating spores. This offers simple high throughput production of transformed plants (Ishizaki et al. 2008).
The genome of M. polymorpha is becoming increasingly well characterised. The Y chromosome of M. polymorpha (10MB)has been sequenced (Yamato et al. 2007), and extensive collections of ESTs have been characterised (2.5 million ESTs total). The ~280MB genome of a Japanese isolate of M. polymorpha (tak1/tak2) is being sequenced currently at the DOE Joint Genome Institute (PIs: John Bowman and Sandra K. Floyd (Monash Univ.), Takayuki Kohchi and Hideya Fukuzawa (Kyoto Univ.), and Kanji Ohyama (Ishikawa Prefectural Univ.)).
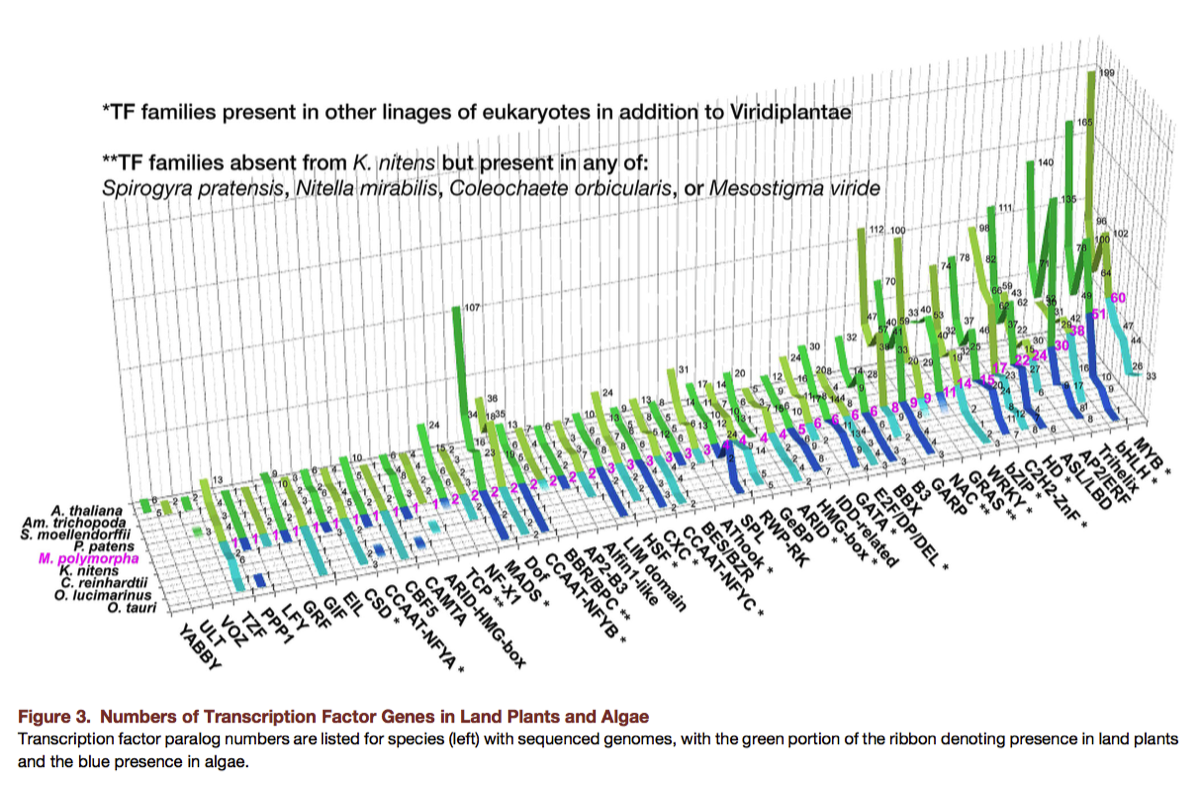
Early indications are that while individual M. polymorpha genes may be longer than counterparts in Arabidopsis, the M. polymorpha genome has a highly streamlined architecture, with smaller gene families and less redundancy compared to higher plants. For example, Sasaki et al. (2007) compared receptor-like kinases (RLKs) in M. polymorpha and Arabidopsis. They identified 600 RLK genes in 52 families in Arabidopsis, compared to 29 RLK genes in 26 families identified in M. polymorpha.
The relative simplicity of genetic networks in liverworts, combined with the growing set of genetic manipulation, culture and microscopy techniques, are set to make these lower plants major new systems for analysis and engineering.
References:
Ishizaki K, Chiyoda S, Yamato KT, Kohchi T. Plant Cell Physiol.49:1084-91, 2008.
Sasaki G, Katoh K, Hirose N, Suga H, Kuma K, Miyata T, Su ZH. Gene. 401:135-44, 2007.
JGI Marchantia Genome sequencing effort:
http://www.jgi.doe.gov/sequencing/why/99191.html
Further information about M. polymorpha can be found at:
(i) The Marchantia Exchange (www.marchantia.org) - based here in Cambridge. The site contains an extensive bibliography with downloadable PDFs, images, useful web links and a forum for technical exchange.
(ii) Marchan Genomics (marchantia.pmb.lif.kyoto-u.ac.jp) - based in Kyoto. The site contains a wide range of very useful information about culture and growth of M. polymorpha and genomic and genetic resources. The site is written in Japanese, but use of Google Translate in your web browser provides a workable solution for foreigners.
Both sites require registration for full access.
Jim Haseloff, University of Cambridge.
Marchantia polymorpha is the best characterised of the liverwort. It is a thalloid liverwort, growing as flat sheet-like tissues which possess distinct upper and lower surfaces. The lower surface of the plant has specialised root-like cells, or rhizoids. The body of the thallus contains oil bodies, present as scattered differentiated cells. This is characteristic of the liverwort group. The upper surface has a modular structure, with repeated formation of units that form primitive cell complexes adapted for photosynthesis and gas exchange. Each unit is marked by a single, permanently open central pore. The liverworts have less efficient systems for water transport and retention than higher plants, and can show marked tolerance to desiccation.
M. polymorpha plants are easy to grow. They show distinctly weed-like properties, growing vigorously on soil, artificial media such as vermiculite and rockwool, and sterile media.
The liverworts have alternate haploid and diploid generations. Like other Bryophytes, the gametophyte or haploid generation is dominant phase of the life cycle. M. polymorpha has a global distribution, and is often found as a weed in horticulture. The plant produces vegetative propagules. These form vegetatively inside conical splash cups. Superficial cells on the inside of a cup undergo cell proliferation to form a group of cells carried on a short stalk, the cells continue to proliferate in regular fashion to form a bilobed gemma (see right). This is eventually detached from the stalk, and can be dispersed from the cup, typically by water splash. The propagules are robust and long-lived, even tolerating desiccation. Gemma can be used for simple vegetative propagation and amplification of plantlets during experiments.
Haploid liverwort plants are either male or female, with sex determined by presence of X or Y chromosomes. A PCR-based assay can be used to determine the sex of M. polymorpha plants. Gametogenesis is under environmental control, and can be induced by exposure to far-red light. Antheriophores and archegoniophores grow upward from the Marchantia thallus, with gamete-forming structures supported by pedestals. Antheridia form on the upper surface of a disc, and archegonia form underneath an arrangement of spokes. Genetic crosses can be performed by transfer of sperm from mature antheridiophores to archegonia by pipette. After fertilisation and zygote formation, the diploid phase of the life cycle continues with cell proliferation, meiosis and spore formation. The spores and elaters are packaged within yellow sporangiophores, suspended under archegoniphores. The elater cells are thickened spiral-shape cells which are hydroscopic and change their helical pitch. These aid the release of spores. Spores can be stored in a cool desiccated state for over a year.
Similar to other lower plants, M. polymorpha regenerates easily, and was used in many early tissue culture studies. A number of plastid and nuclear transformation techniques have been developed for M. polymorpha, including a recent method for Agrobacterium tumefaciens mediated transformation using germinating spores. This offers simple high throughput production of transformed plants (Ishizaki et al. 2008).
The genome of M. polymorpha is becoming increasingly well characterised. The Y chromosome of M. polymorpha (10MB)has been sequenced (Yamato et al. 2007), and extensive collections of ESTs have been characterised (2.5 million ESTs total). The ~280MB genome of a Japanese isolate of M. polymorpha (tak1/tak2) is being sequenced currently at the DOE Joint Genome Institute (PIs: John Bowman and Sandra K. Floyd (Monash Univ.), Takayuki Kohchi and Hideya Fukuzawa (Kyoto Univ.), and Kanji Ohyama (Ishikawa Prefectural Univ.)).
Early indications are that while individual M. polymorpha genes may be longer than counterparts in Arabidopsis, the M. polymorpha genome has a highly streamlined architecture, with smaller gene families and less redundancy compared to higher plants. For example, Sasaki et al. (2007) compared receptor-like kinases (RLKs) in M. polymorpha and Arabidopsis. They identified 600 RLK genes in 52 families in Arabidopsis, compared to 29 RLK genes in 26 families identified in M. polymorpha.
The relative simplicity of genetic networks in liverworts, combined with the growing set of genetic manipulation, culture and microscopy techniques, are set to make these lower plants major new systems for analysis and engineering.
References:
Ishizaki K, Chiyoda S, Yamato KT, Kohchi T. Plant Cell Physiol.49:1084-91, 2008.
Sasaki G, Katoh K, Hirose N, Suga H, Kuma K, Miyata T, Su ZH. Gene. 401:135-44, 2007.
JGI Marchantia Genome sequencing effort:
http://www.jgi.doe.gov/sequencing/why/99191.html
Further information about M. polymorpha can be found at:
(i) The Marchantia Exchange (www.marchantia.org) - based here in Cambridge. The site contains an extensive bibliography with downloadable PDFs, images, useful web links and a forum for technical exchange.
(ii) Marchan Genomics (marchantia.pmb.lif.kyoto-u.ac.jp) - based in Kyoto. The site contains a wide range of very useful information about culture and growth of M. polymorpha and genomic and genetic resources. The site is written in Japanese, but use of Google Translate in your web browser provides a workable solution for foreigners.
Both sites require registration for full access.
Jim Haseloff, University of Cambridge.
Marchantia polymorpha genome published
Publication of characterisation of the Marchantia polymorpha genome - with description of the streamlined genome of this key lower plant, with low regulatory gene counts and evolutionary stepping-stone between algae and higher plants. A great example of selfless community building by John Bowman, Takayuki Kochi, Tom Yamato and many colleagues!
Bowman et al. Cell 171:287–304 (2017). Click to download PDF
Bowman et al. Cell 171:287–304 (2017). Click to download PDF
Supplemental material from our group
Analysis of Cambridge isolates of Marchantia polymorpha, Bernardo Pollak, Mihails Delmans and Jim Haseloff. Summary of comparison of Cam-1, Cam-2 and Tak-1 isolates, and transcriptomic analysis of germinating sporelings. Cambridge contribution to Bowman et al. Cell 171:287–304 (2017). Click to download PDF
MarpoDB: An open registry for Marchantia polymorpha genetic parts
MarpoDB is an open source database for Marchantia genes and DNA parts, which has been designed as a tool for a synthetic biology workflow. Among its features are precompiled cross-database querying to InterPro, Pfam signatures and non redundant Viridiplantae BLAST annotations; BLAST querying to Marchantia genes; sequence export in GenBank format; recoding of sequences to the common syntax for type IIS assembly and exchange of DNA parts; and a user interface for gene models and sequence exploration. MarpoDB is a platform for plant synthetic biology experiments in this model system. Mihails Delmans*, Bernardo Pollak* and Jim Haseloff, MarpoDB: An Open Registry for Marchantia polymorpha Genetic Parts, Plant Cell Physiol. 58: e5(1–9) (2016). Click to download PDF.
(Additional resources: GitHub site for the source code and URL for the active database)
(Additional resources: GitHub site for the source code and URL for the active database)

Above. Marchantia polymorpha plant (photo: Jim Haseloff)

Above. Upper surface of M. polymorpha thallus, showing air pores (photo: Jim Haseloff)
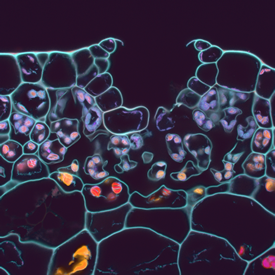
Above. Cross section of M. polymorpha thallus, showing air pore and photosynthetic tissues (confocal microscopy: Jim Haseloff)

Above. Splash cup and gemmae on M. polymorpha thallus(photo: Jim Haseloff)

Above. Development of gemmae in M. polymorpha (The Hepaticae and Antherocerotae of North America, Rudolf Schuster, 1966)
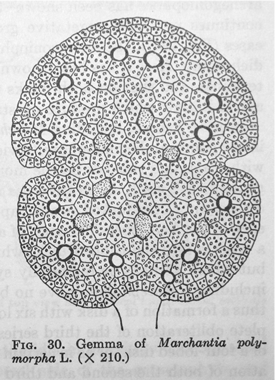
Above. Structure of a gemma in M. polymorpha (Cryptogamic Botany Vol II, Gilbert Smith, 1955)

Above. Formation of spore capsules in M. polymorpha (photo: Jim Haseloff)


Above. Formation of young M. polymorpha plants in axenic culture after germination of surface sterilised spores (photo: Jim Haseloff)

Above. Comparison of RLK gene families in Arabidopsis and M. polymorpha (Sasaki et al. 2007)